The Circulatory System
Mammals need mass transport systems
Organisms need oxygen and glucose to produce energy in aerobic respiration. Small organisms, like bacteria, can obtain these by diffusion because of the short diffusion distance. But in multicellular organisms, the diffusion distance is too large and diffusion would be too slow to reach the cells in the centre of their body, so they have mass transport systems.
Mass transport systems, such as the circulatory system in mammals, deliver oxygen and glucose to all body cells. They also remove waste products (carbon dioxide and water).
Types of circulatory system
Circulatory systems can be open or closed. Closed circulation can either be a single or double system:
Open circulatory systems – not enclosed in blood vessels. Insects have an open circulation – blood flows out of the heart and into the body cavity. Blood flows around the insect’s organs, gradually directed back to the heart through a series of valves.
Closed circulatory systems - blood is enclosed inside blood vessels. All vertebrates (i.e. mammals and fish) have a closed circulation. Arteries take blood away from the heart and towards organs. Substances move between the bloodstream and tissues at the capillaries. Veins take blood from tissues back to the heart.
Single circulation – blood travels in one loop, passing through the heart once for each complete circuit of the body. In fish, the heart pumps blood to the gills, then straight to the body tissues, before returning back to the heart.
Double circulation – blood travels in two loops, from the heart to the lungs for gas exchanges, then back to the heart before being pumped around the body. Mammals have this type of circulation. The two loops are known as the pulmonary system (to the lungs) and the systemic system (to the body).
Structure of blood vessels
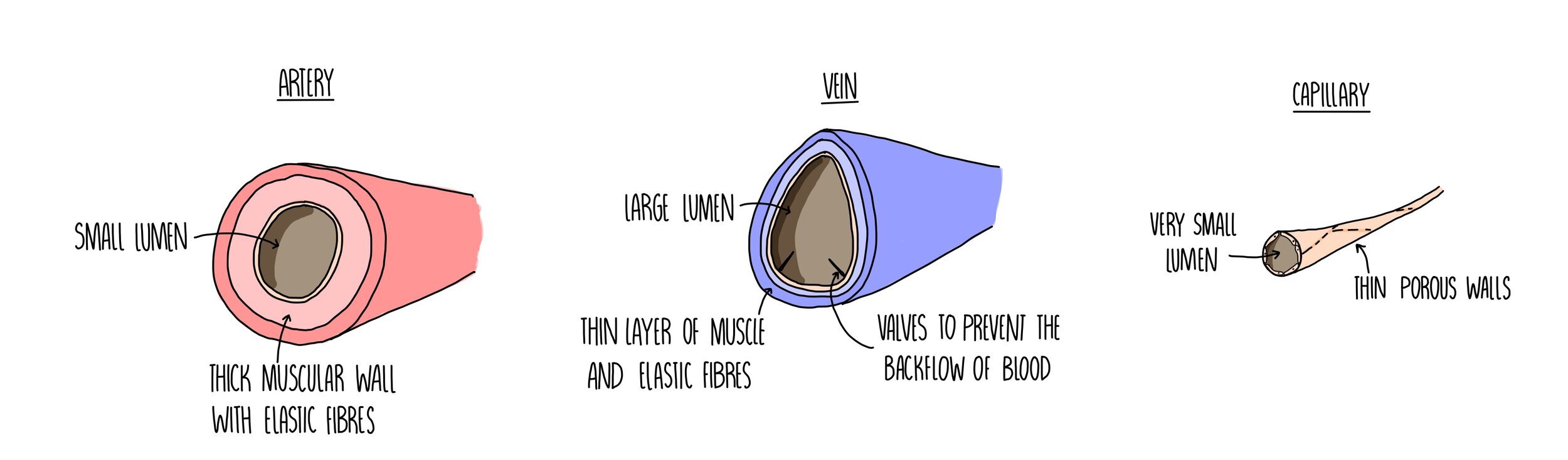
Arteries
Carry blood away from the heart to the various organs of the body.
They need to cope with the high pressure generated from the heart forcing out blood with each heartbeat. This is why they have a really thick muscular wall containing lots of elastic tissue.
The inner lining of the arteries, called the endothelium, is folded which allows the artery to expand (elastic recoil) which also helps it to withstand the high pressure.
The small lumen ensures a high pressure is maintained.
Veins
Carry blood from the organs of the body towards the heart.
Blood is flowing at a much lower pressure so veins have a large lumen and much thinner walls containing little elastic fibres or muscle tissue.
Valves prevent the slow-moving blood from flowing backwards.
The contraction of nearby body muscles helps blood to flow through veins.
Capillaries
Connect arteries and veins.
Substances move out of the blood to the body tissues - things like oxygen, glucose and mineral ions. Any waste products, such as carbon dioxide and water, will move out of the body tissues and into the capillaries.
Small holes (pores) enable the exchange of substances.
Walls are just one cell thick which reduces the diffusion distance for these substances.
Tissue fluid
Tissue fluid is the fluid that bathes cells in tissues. It is made up of the substances that are small enough to move out of the capillary, such as water, oxygen, glucose and mineral ions. It also contains the waste products released from cells, like carbon dioxide, water and urea. It doesn’t contain things that are too big to be forced out of the capillary, so there are no red blood cells or large proteins.
The formation of tissue fluid takes place when smaller substances are pushed out of the capillary bed:
At the arteriole end of the capillary, hydrostatic pressure in the capillary exceeds that of the tissue fluid. This forces fluid and dissolved substances out of the capillary (down a pressure gradient) and into the spaces around the cells, forming the tissue fluid.
At the venule end of the capillary, there is less water (because it moved out of the capillary at the arteriole end). This means that the water potential is lower inside the capillary compared to the tissue fluid. Water moves back into the capillary by osmosis down a water potential gradient.
Excess tissue fluid drains into the lymphatic system which eventually returns the fluid back into the bloodstream.

Heart structure
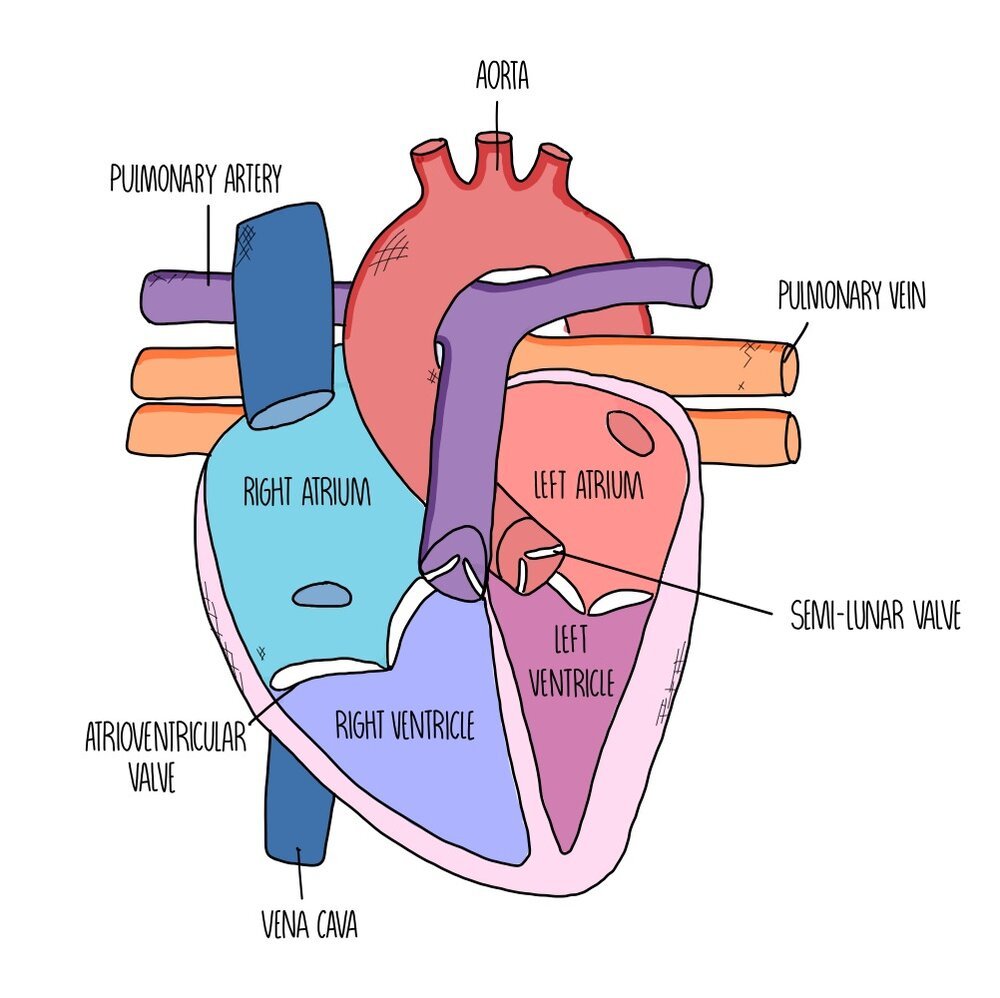
The heart is made up of four chambers divided into two sides. The left side of the heart has a thicker wall, as it needs to pump more strongly to deliver blood all around the body (whereas the right side just needs to send the blood to the lungs). The left side carries oxygenated blood whereas the right side carries deoxygenated blood.
The chambers at the top are called atria and these chambers receive the blood from the veins supplying the heart. Blood flows from the atria to the ventricles, which are separated from the atria by atrioventricular valves to prevent blood flowing in the opposite direction. There are another set of valves between the ventricles and the arteries which are called the semi-lunar valves as they look like little half-moons.
The main artery which takes oxygenated blood from the left side of the heart to the rest of the body is called the aorta whereas the artery which delivers deoxygenated blood between the right side of the heart and the lungs is called the pulmonary artery. The major vein which returns blood from the body to the right side of the heart is the vena cava and the vein which ferries blood from the lungs to the heart is called the pulmonary vein.
The heart muscle itself also needs its own blood supply, so that it can get plenty of oxygen and glucose to keep respiring and keep pumping - these are called the coronary arteries. It’s a blockage in these coronary arteries which leads to a heart attack.
The Cardiac Cycle
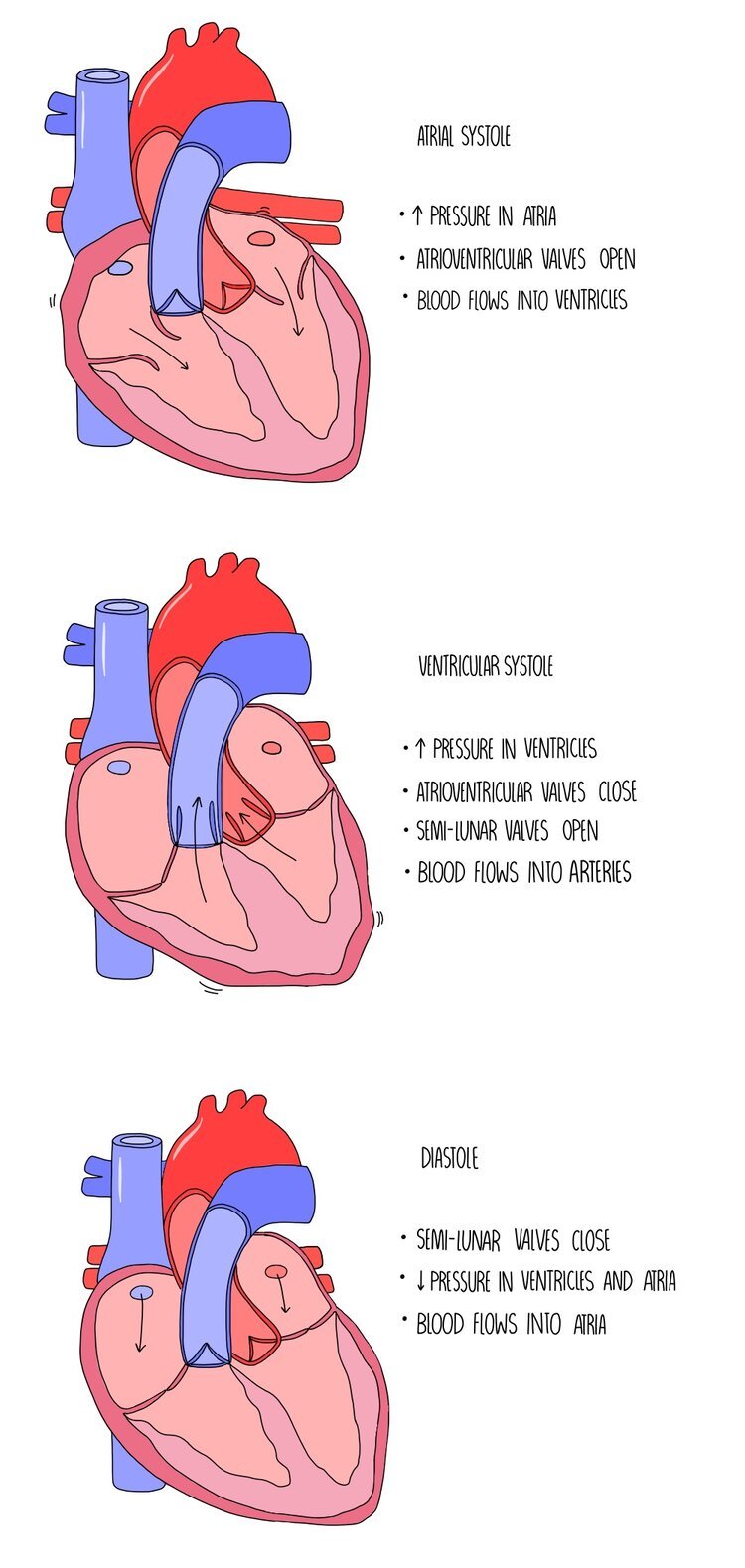
The cardiac cycle is a coordinated sequence of contractions and relaxations by the heart muscle which causes blood to move from the atria, into the ventricles and then the arteries. Muscle contractions are referred to as systole and relaxation is referred to as diastole. The movement of blood through the different compartments of the heart rely on changes in volume and pressure as a result of the contraction of the heart muscle walls. It occurs in the three following stages:
1. Atrial systole
The atria contract while the ventricles relax. This decreases the volume inside the atria which increases the pressure. This increased pressure forces the atrioventricular valves open and pushes blood into the ventricles.
2. Ventricular systole
The ventricles contract while the atria are relaxed. This decreases the volume inside the ventricles which increases the pressure. The pressure is higher in the ventricles compared to the atria, which forces the atrioventricular valves closed and causes the semi-lunar valves to open. The closure of the AV valve is important because it prevents the back-flow of blood into the atria. Blood is forced out of the ventricles and into the arteries (the aorta and the pulmonary artery).
3. Diastole
Both the atria and ventricles are relaxed so the pressure is low in both chambers. Since the pressure is higher in the arteries than in the heart chambers, the semi-lunar valves are forced closed which prevents blood flowing backwards into the ventricles. Blood is returned to the heart and the atria fill with blood.

Myogenic contraction

Cardiac muscle is myogenic, which means that it can contract on its own accord without being stimulated by nerve impulses. It is coordinated by a nodule in the wall of the right atrium called the sino-atrial node (SAN). The SAN is referred to as the ‘pacemaker’ of the heart because it sets the rhythm of heart contractions by creating regular waves of electrical activity which spread out to the atrial walls. This wave of electrical activity causes the right and left atria to contract at the same time. There is a band of non-conducting tissue between the atria and the ventricles which prevents the wave of electricity from reaching the ventricles. The electrical wave is instead directed towards the atrioventricular node (AVN) which is located between the atria and the ventricles. The AVN passes the wave of electrical activity onto a group of muscle fibres located in the centre of the heart, called the bundle of His. There is a delay before the wave of excitation passes onto the bundle of His to ensure that the ventricles only contract after the atria have finished emptying with blood. The bundle of His then passes on the wave of electrical activity to the Purkyne fibres, found along the bottom of the heart around the ventricular walls. This causes the left and right ventricles to contract simultaneously, from the bottom up.
Electrocardiographs
An electrocardiograph is a machine which can record the electrical activity of the heart. Whenever a part of the heart contracts, the membrane of muscle cells becomes depolarised (it loses charge) and repolarises (gains charge) when the muscle relaxes. This change in charge in the heart muscle can be detected by an electrocardiograph, which uses electrodes placed against the patient’s chest. The change in electrical activity is displayed as an electrocardiogram (ECG) which can be used by doctors to diagnose heart problems.
The first, small bump on an ECG is called the P wave which is caused by the atria contracting. The large, sharp spike is called the QRS complex and is caused by the ventricular contraction. The final bump is referred to as the T wave and results from repolarisation of the ventricles as they relax. You can pick out any of these points on the ECG to measure the time taken for one heart beat to occur - e.g. the time taken from one P wave to the next P wave. The height of the wave is proportional to the strength of contraction - the higher the wave, the greater the depolarisation of the muscle cell and the stronger the contraction.

ECG can be used to diagnose heart problems such as tachycardia (abnormally fast heart rate), bradycardia (abnormally slow heart rate), ectopic heartbeats (‘extra’ heartbeats) and fibrillation (complete loss of rhythm). The image below shows how the ECG from patients with some of these heart problems would look. Abnormal heart rates or fibrillation may be a consequence of cardiovascular disease or other heart conditions, such as damage to the heart muscle.

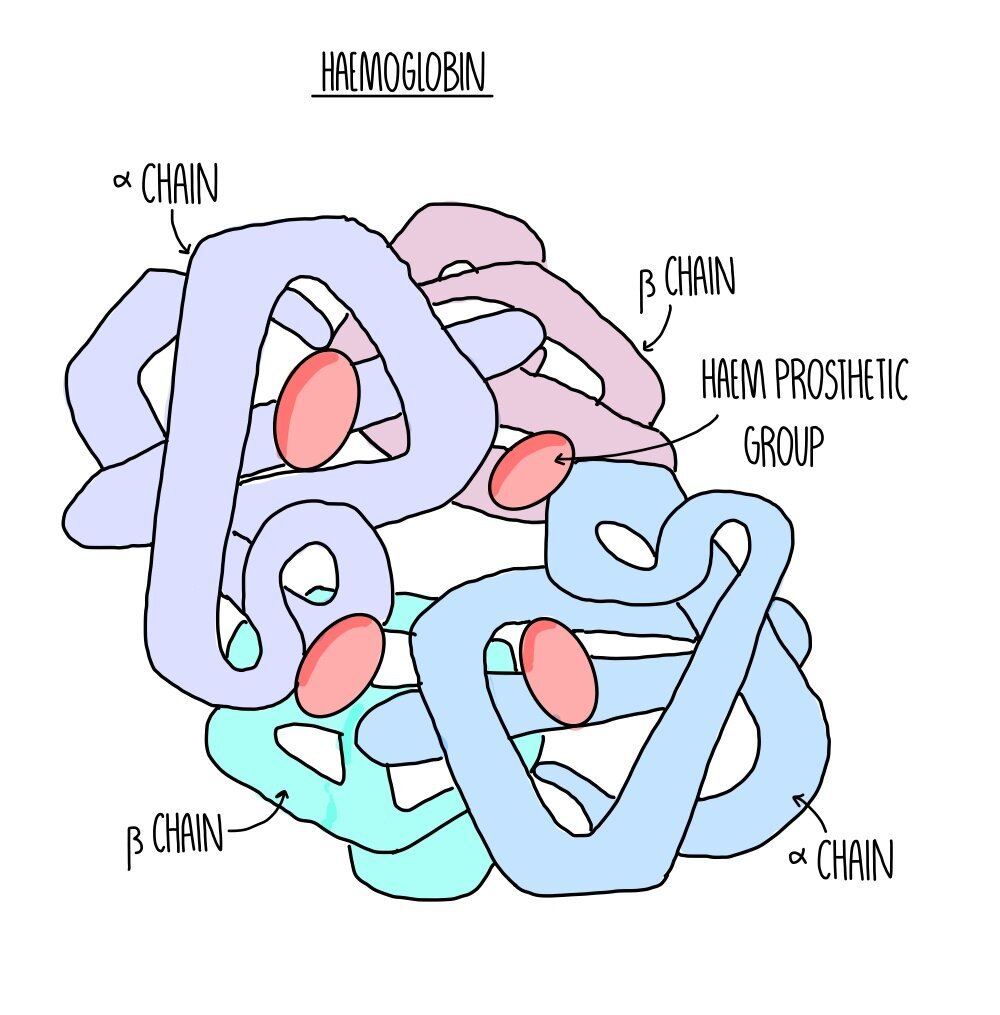
Haemoglobin
Oxygen is transported by binding to haemoglobin in red blood cells. Haemoglobin is a large, globular protein with a quaternary structure made up of four polypeptide chains. Its four subunits each possess a haem group containing an iron ion. Four oxygen molecules can bind to the four haem groups to form oxyhaemoglobin.
Haemoglobin saturation and dissociation curves
Haemoglobin has different affinities for oxygen at different oxygen concentrations (partial pressures). Affinity means how ‘sticky’ haemoglobin is – the higher it’s affinity, the stronger it binds. Haemoglobin binds to oxygen with high affinity in the lungs, where there is a high partial pressure of oxygen. Haemoglobin binds with low affinity at respiring tissues, which there is a low partial pressure of oxygen. This causes oxyhaemoglobin to deposit oxygen at the tissues so they can use it for aerobic respiration.

You can see this on a dissociation curve, which shows how saturated haemoglobin is with oxygen at different partial pressures.
The graph has a sigmoid, or S-shaped, because binding of the first oxygen molecule causes oxygen to change shape and uncover the other oxygen binding sites. This makes it easier for further oxygen molecules to bind and is known as cooperative binding. Once three oxygen molecules are bound, it is harder for the final one to bind because there is only one binding site available.
Fetal haemoglobin
Fetal haemoglobin has a higher affinity for oxygen compared to adult haemoglobin. This means that it binds more strongly, holding on to the oxygen as it makes its way around the mother’s body. It ensures that it is still in the oxyhaemoglobin form by the time It gets past the placenta to reach the fetus. If fetal haemoglobin had the same affinity for oxygen as adult haemoglobin its blood wouldn’t be saturated enough. On a dissociation curve, fetal haemoglobin lies slightly to the left.
The Bohr Effect

Haemoglobin’s affinity for oxygen also changes at different concentrations of carbon dioxide. At higher partial pressures of carbon dioxide, haemoglobin has a lower affinity for oxygen, causing oxyhaemoglobin to dissociate more readily.
This is useful because tissues with high rates of respiration (so greater need for oxygen) will be producing larger amounts of carbon dioxide. This ensures that oxygen is unloaded from haemoglobin at respiring tissues.
High carbon dioxide concentration shifts the dissociation curve to the right. It is referred to as the Bohr Effect.
The Bohr effect occurs due to the chloride shift:
Carbon dioxide released from respiring cells moves into red blood cells and reacts with water to form a weak acid called carbonic acid.
Carbonic acid dissociates to form a hydrogen ion and hydrogencarbonate ion.
The increased acidity causes oxyhaemoglobin to unload its oxygen. Hydrogen ions bind to haemoglobin, forming haemoglobonic acid.
The hydrogencarbonate ions diffuse out of the red blood cell, transported back to the lungs in the blood plasma.
To balance the charges, chloride ions diffuse into the red blood cell – this is called the chloride shift.
When the blood reaches the lungs, the hydrogen ion recombines with the hydrogencarbonate ion to form carbon dioxide and water. The carbon dioxide diffuses into the alveoli and is breathed out.